ABSTRACT
“Theranostics” refers to therapy and diagnosis. This paper presents a new method for therapy or diagnosis (or both) for malignant tumors. The underlying premise is that cancer cells with their supernumerary centrioles have a significantly higher electromagnetic field than that of normal cells. This enhanced field can then serves as a “biomarker” or target for the attraction of nanoparticles - specifically: Super Paramagnetic Nanoparticles (SPIONS) which can provide the desired theranostics. Divided into nine sections, the paper provides the bases for these assertions.
Introduction
Perhaps the greatest challenge for nanotechnologists is the development of nanoparticles for finding and destroying cancer cells. In this paper we review efforts toward this end. We then propose a new method for accomplishing this.
The paper provides an elaboration of a poster presented at Herald International Conference on Nanomedicine & Nanotechnology [1].
The premise is that cancerous tissue has a greater electromagnetic field surrounding it than that of normal tissue. This enhanced field can then be a target biomarker for charged nanoparticles.
As early as 1969 Paul Schafer claimed that disrupted centriole geometry is found in cancer cells [2-4]. He also showed experimentally that large magnetic fields can distort centriole geometry (Centrioles are small perpendicular cylindrical structures lying adjacent to the nucleus in human and animal cells).
Unfortunately Schafer’s findings received relatively little attention since it was apparently overshadowed by the discovery and interpretation by Watson and Krick at about the same time, of the double helix within the cell nucleus [5]. But then, at about the turn of this 21st century, biological researchers began to look again at disrupted centrioles and clusters of centrioles as cancer markers. References 6-20 provide a sampling of these many investigations.
There remain, however, several significant gaps in nanomedicine and cancer therapy. The greatest of these is the absence of a reliable method for safely delivering nanoparticles to cancer cells without harming normal cells. Also the toxicity of nanoparticles remaining in a body has not been fully investigated. Finally, application with brain tumors presents a special challenge due to the blood-brain barrier.
In a series of recent papers AV Singh, et al., summarize current issues in nanomedicine and cancer therapy [21-25]. They also discuss the gaps in the use of nano particles for cancer therapeutics [25-27].
Centrioles
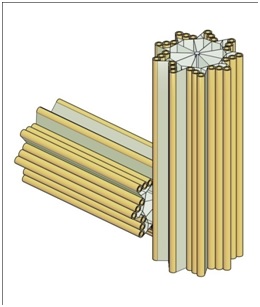
The interior structure of eukaryotic (human and animal) cells is well known. For our purposes, for tumorigenesis, we focus on centrioles - a small pair of perpendicular cylinders adjacent to the nucleus. Errors in centriole geometry are believed to be a source of tumorigenesis [28].
The centrioles are distinctly different from all other organelles and organs:
1) They have precise geometry
2) They have no membrane cover
3) They may be viewed as hollow cylinders with nine radial blades, each having themselves three hollow cylinders known as “microtubules”, forming the centriole circumference
4) Viewed as a cylinder, each centrioles is approximately 400 to 500 nm long and 200 nm in diameter
5) Centrioles occur in pairs (often called the “mother” and “daughter”) with the daughter being perpendicular to the mother and attached to the mother at the base
6) The centrioles lie in a cloud of many proteins which together with the centrioles is known as the “centrosome”
Figure 1 provides a sketch of a typical centriole pair.
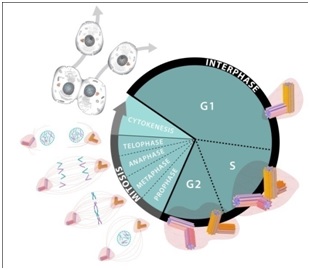
The centrioles play a central role in cell division. Although this process is generally well understood it may be helpful to some readers to briefly review the cell cycle (birth to separation) [29-33]. The cycle may be viewed in two phases: the “M-phase” (mitosis) and the “interphase” as represented in figure 2.
The M-phase consists of several subphases called: “Prophase”; “Metaphase”; “Anaphase”; “Telophase”; and “Cytokinesis”. This set of subphases is also known as “mitosis”. Curing mitosis the nucleus is divided and the chromosomes are separated. The cytoplasms with its various organelles are also separated into two halves, each half following one of the nucleus halves.
The Interphase, which is considerably longer than the M-phase also consists of several phases: G1 (or “Gap1”) for growth; S-phase, for more growth and DNA duplication; and G2 (or “Gap2”) for still more growth and for preparation for division.
During the S-phase the centrioles are also duplicated into two pairs, and then during prophase the centriole pairs are separated with the newer pair moving to the opposite side of the nucleus. The centriole pairs now on opposite sides of the nucleus extend their microtubules to form the mitotic spindle, and then pull apart the nucleus [29-33].
As noted previously the centrioles are composed of nine “blades” of Microtubules (MTs) with each blade having three MTs. Thus each centriole has a total of 27 parallel MTs, and some centrioles have an additional two MTs along their axes.
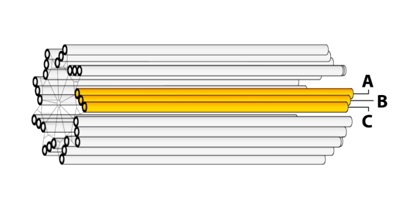
For a typical blade of MTs, each MT does not have the same length: The interior MT (closest to the centriole axis) is the longest. The exterior MT (closest to the centriole perimeter) is the shortest. The third MT then has intermediate length. They are labeled: “A”, “B”, and “C” as represented in figure 3.
New (“daughter”) centrioles are grown from the side at the base of the “mother” centriole, with the axis of the daughter being perpendicular to the axis of the mother. The daughter growth begins on the exterior side of one of the C-MTs.
With there being nine C-MTs, it is not clear which is to be selected for the base of the daughter centriole.
The growing process is believed to occur as follows: During the S-phase of the cell cycle, as the DNA is being separated, a small quantity of Asterless (AsP) is deposed at the base of the selected C-MT. (AsP is the orthologue of Cep152 [34]. The AsP/Cep152 then recruits a patch of the polo-like kinase enzyme: Plk4 which then in turn recruits the protein: SAS-6 for the base of the ensuing daughter centriole.
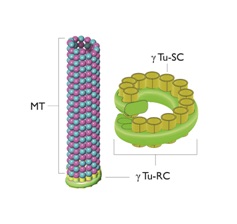
SAS-6 projects nine outward spokes whose ends attract three spots of Gamma (ϒ) tubulin. The ϒ-Tubulin Ring Complex (ϒTu-RC) and the ϒ-Tubulin Small Complex (ϒTu-SC) [7,35-43].
Figure 4 provides a simplified representation of this process.
Microtubules
As noted earlier the centrioles are immersed in a cloud of many proteins with the system of the centrioles and proteins known as the “centrosome”. The centrosome is also known as the Microtubule Organizing Center (MTOC) since the proteins making up the microtubules come out of the centrosome via the ϒTu-SC.
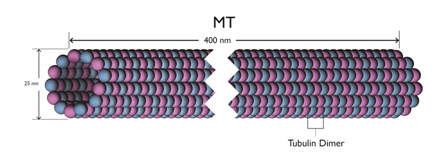
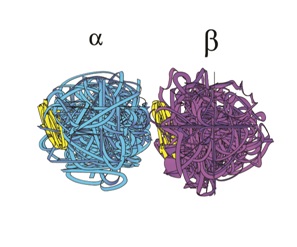
These Microtubules (MTs) are themselves long hollow cylinders - approximately 400 nm long, 25 nm in outside diameter and 15 nm in inside diameter. They are composed of longitudinal connected tubulin dimers. A tubulin dimer is a connected Alpha (α) and Beta (β) tubulin.
Figures 4 and 5 provide sketches of MT and figure 6 shows a sketch of a tubulin dimer.
Around the circumference of MT there are 13 filaments. This odd number shows that there is no distinct or uniform pattern of α and β tubulin around the circumference. This in turn implies that the filaments can move longitudinally relative to the overall MT structure.
At the distal end of a MT, if a filament is moving in the distal direction, the end α-β dimers may fall away into the cytoplasm. This activity is known as “catastrophe”, and the length of the MT could be shortened. Alternatively, if the filament is moving in the distal direction and the dimers at the distal end do not fall into the cytoplasm, the MT is lengthened. This is known as “rescue”.
In addition to both catastrophe and rescue, the MT filaments can also oscillate longitudinally.
Reference 44 provides a more detailed review of microtubules.
Electromagnetics (EMs)
The α-β tubulin dimers have a negative-positive charge – similar to that of a flashlight battery. A moving charge creates an Electromagnetic (EM) field. A Rapid movement such as a MT filament oscillation can create EM fields. With 27 MTs in each centriole, and with each MT having 13 filaments, there are a total of 351 filaments per centriole, and thus 702 filaments for each centriole pair. The oscillations of this large number of filaments can create strong, varying EM fields.
References 45-49 provide a more detailed review of cellular electromagnetics.
Development of an Electromagnetic Biomarker
In a normal cell there is only one pair of centrioles, except during mitosis when there is two pair. In cancer cells, however, there are numerous (or “supernumerary”) centrioles due to errors in the centriole duplication process [7-20]. While there may be many reasons for the duplication going awry (carcinogens, viruses, radiation) a typical error is a “flowering” centriole as suggested in figure 7.
When there are supernumerary centrioles, they tend to cluster together as an enlarged centrosome. This clustering with the resulting thousands of MT filaments will create an enhanced EM field far greater than that of normal cells. This enhanced EM field is then a biomarker distinguishing cancer cells from normal cells. Consequently, this EM biomarker can serve as a target for imaging and therapy. Specifically, in our analysis this EM biomarker is thought to be an ideally suited attractor for magnetically charged nanoparticles.
Nano Theranostics
With theranostics referring to both diagnostics and therapy, if we can get Nanoparticles (NPs) within the cancer cell membrane while at the same time avoiding normal cells, we can both diagnose the extent of the cancerous tumor and also provide therapy to destroy the tumor. Specifically, if the NPs contain metals, such as gold or iron, they can be imaged by x-rays, and thus expose the extent of the tumor. Similarly, if the cell-inserted NPs contain toxic chemicals, the chemical can be released via external control.
If a tumor can be illuminated it can be treated by focused x-rays and/or proton beams. Alternatively, in addition to depositing toxins, magnetically charged NPs can be exposed to oscillating external magnetic fields causing the MTs to vibrate and thermally destroy the cancer cells. In either event the objective is to get NPs inside the cancer cell.
If the NPs are sufficiently small (<20 nm) they will be able to pass through the cell membrane. If the NPs are also magnetic, they will be attracted to the centriole cluster of the cancer cell. The task then becomes: How can we get the NPs sufficiently close to the cancer cells, so the enhanced electromagnetic field of the cancer cell will pull the magnetized NP into the cell? To answer this question, consider first that if the NPs are simply inserted into the blood stream, analogous to chemotherapy, they may be excreted before reaching the tumor or as with chemotherapy the enhanced blood pressure in the vasculature supplying the tumor may tend to push the MTs away from the tumor. Moreover, if the MTs are magnetic, they will be attracted to each other forming clumps which are too large to penetrate the cell membrane, and/or with reduced magnetism to be strongly attracted by the clustered centrioles.
Therefore, what is needed is a direct focus of NPs on the tumor (analogous to the current use of proton beams) but without the NPs clustering together, and also without harm to nearby normal tissue.
To avoid the clustering we can use super paramagnetic NPs which have no magnetic attraction until they are in the presence of a relatively stronger Electromagnetic (EM) field. NPs with iron oxide constituents have been shown to have such properties and are known as “Super paramagnetic Iron Oxide Nanoparticles“(SPIONS) [50-54].
As of this writing the best candidates for development of effective SPIONS are carbon nanotubes with iron oxide cores, known as: “carbon nanoworms”, and similarly carbon nano-spherical shells, known as: “carbon nanospheres”.
Discussion
In addition to SPIONS (e.g., carbon nanoworms and carbon nanopsheres) numerous other nanoparticles have been proposed for tumor treatment, with some success. These include [55-58]:
1) Dendrimers - branching tree-like structures
2) Micelles - spherical, globular structures, with a hydrophobic central core
3) Nanocapsules - fluid filled pouches
4) Fullerenes - carbon bucky balls
5) Liposomes - closed fluid filled spheres with covering layers
6) Polymeric nanoparticles - structures of polymers
7) Ceramic nanoparticles - silicon and other ceramics
8) Metallic nanoparticles - gold, steel, and iron particles
9) Poly(ethylene glycol) nanoparticles - aka “PEGs”
10) SWCNTS - Single-Walled Carbon Nanotubes
Gidwani and Singh have reviewed the possibility and opportunities of using nanoparticles to cross the blood brain barrier [59]. Singh et al., also consider the use of nanoparticles in treating multiple sclerosis and the safety of using carbon nanoparticles in theranostic applications [60,61].
Recently it has been discovered that cancer tumors possess an excess of the vitamin: folic acid [62-65]. Thus, as with an enhanced Electromagnetic (EM) field folic acid is also a biomarker. PEGs (see 9 above) are believed to be attracted to folic acid receptors.
Chemotherapy is a commonly used therapy for cancer. A difficulty with toxic chemicals injected into the blood stream is that the vasculature supplying a tumor is often enlarged with high back pressure. This high pressure can reduce the amount of chemical entering the tumor so that all cells of the tumor are not necessarily reached by the chemical. Over time these unreached cells may cause the tumor to recur. Also, the toxic chemicals are harmful to normal cells.
A principal advantage of nanoparticle therapy is that the nanoparticles can be directed toward the cancer tumor without harmful effects on the surrounding normal tissue with its lower electromagnetic attraction. (Nanoparticles injected into the blood stream are subjected to the same high pressure resistance by the tumor vasculature as the chemical toxins).
Finally although the number of cancer cells in a tumor is large, there is practically no limit to the number of nanoparticles which can be directed toward the tumor’s cancer cells.
The future work is thus well-defined: We need to improve the efficacy of the nanoparticles - particularly the SPIONS and the PEGs.
Acknowledgement
Support for this research has been provided by the Schafer Foundation for Centriolar Research. The encouragement of Bardyl Tirana and Roger Adelman is also acknowledged, and the assistance of Charlotte Better and Mary Schartman in preparing the manuscript is appreciated. Finally the helpful suggestions of the reviewers are appreciated.
Figures
Figure 1: A Typical centriole pair and their centriolar blades.
Figure 2: Cell cycle.
Figure 3: Centriole blades.
Figure 4: A microtubule, the Gamma-Tubulin Ring Complex (ϒTu-RC) and the Gamma-Tubulin Small Complexes (ϒTu-SCs).
Figure 5: A microtubule length.
Figure 6: A Tubulin Dimer.
Figure 7: A flowering centriole.